يقول داروين في كتابه أصل الأنواع: "إذا كان من الممكن إثبات وجود أي عضو معقد، لا يُرجَّح أنه قد تشكل عن طريق العديد من التعديلات المتعاقبة والطفيفة، فسوف تنهار نظريتي تماما".(1)
يعتبر بعض المتحمسين لداروين مقولته السابقة دعوة للتحدي، في حين يرجعها المتابعون إلى الشك وعدم الثقة المتأصلان في نظريته التي تناقش أصل الأنواع. أيًّا كان المقصد، فأطروحتنا التي بين أيديكم اليوم هي استجابة لدعوة داروين للتحدي، أو تأكيدا لشكه.
• ما هي الحياة، وكيف تفسر الداروينية تعقيدها؟
• ما هي تلك النظم الحيوية غير القابلة للاختزال، وكيف تمعن في تحدي الداروينية؟
• هل تفوز تلك النظم في التحدي الذي وضعه داروين وتحقق توقعه بانهيار نظريته، أم تؤكد أن الداروينية ما زالت ثابتة رغم التحديات؟
• كيف تعاطى أنصار الداروينية مع تلك المعضلة، وهل استطاع كينيث ميلر تخطيها وتفنيدها كما ادعى خلال شهادته في محاكمة التصميم الذكي بمنطقة دوڤر؟
• هل الصراع حول تفسير الحياة بشكل عام، تحسمه المحكمة؟ أم أن المحاكمة ستضطرنا لإعادة المحاكمة؟
هنا نحاول الإجابة على هذه الأسئلة، فتابعوا معنا إعادة المحاكمة.
- ما هي الحياة؟
هي الفرق بين الحي والميت، الحياة والجماد. هكذا ظل منتهى علمنا بالحياة حبيسًا داخل حدود هذا التوصيف، فالحياة هي اللغز الأعظم الذي حير الفلاسفة وأعجز البيولوجيون وأعيا أجيالا متعاقبة من الباحثين.
وحتى في الدين، فالروح هي الكينونة المبهمة بداخل كل مخلوق حي، تميزه عن الجماد، وتفارق بدنه عند الموت. سألت عنها اليهود رسول الله ﷺ، فأجاب إجابة واضحة مباشرة، بما أوحى إليه ربنا سبحانه وتعالى: (وَيَسْأَلُونَكَ عَنِ الرُّوحِ قُلِ الرُّوحُ مِنْ أَمْرِ رَبِّي وَمَا أُوتِيتُمْ مِنَ الْعِلْمِ إِلَّا قَلِيلًا) [الإسراء: 85].
منذ بدأ الثورة العلمية، كان هناك نزاع مستمر تدور رحاه في أوساط المعرفة البيولوجية حول توصيف الحياة ومصدريتها، انقسمت فيه الآراء إلى معسكرين: أحدهما يعلن أن الكائنات الحية ليست مختلفة إطلاقا عن المادة غير الحية، وكانوا يدعون أنفسهم أحيانا بالآليين Mechanists، أو الفيزيقيين Physicalists. وعلى الجانب المضاد يقف معسكر الحياتيين Vitalists، وكانوا ينادون برأي مفاده أن للكائنات الحية خصائص لا يمكن وجودها في المادة الخاملة، وأن المفاهيم البيولوجية لا يمكن أن تكون خاضعة لقوانين الفيزياء والكيمياء. أسست الحياتية منهجا راسخا بداية من القرن السابع عشر، وشكلت عقبة قوية في وجه الفلسفة الآلية المادية، التي ادعت أن الحيوان ما هو إلا آلة، وأن كل مظاهر الحياة هي عبارة عن مادة في حالة حركة. وتبنت الحياتية فكرة وجود مادة خاصة وقوة حيوية غير موجودة في الجمادات هي ما تميز الحياة، وكان كثير من أتباع تلك المدرسة من الغائيين Teleologists الذين يعتقدون أن الحياة وجدت لتحقيق غاية.
تسبب تشبث الحياتية بوجود تلك القوة الحيوية المبهمة في التعجيل بسقوطها؛ على اعتبار أنها فكرة ميتافيزيقية أكثر منها علمية، حتى انزوت تقريبا عن المشهد العلمي مع بدايات القرن العشرين وصعود الفلسفة المادية والنهج العلماني في تناول البيولوجيا.
 في أواخر القرن التاسع عشر أتت الداروينية حاملة في كنفها نزعة مادية عاتية، كمخرج من سطوة الفكر الديني الأنجليكاني الذي تسيد المشهد البيولوجي آنذاك، فأنكرت وجود الغاية الكونية، واستبدلتها بمفهوم الانتخاب الطبيعي لقيادة التنوع الأحيائي، من خلال مبدأ الصراع من أجل البقاء، بالإضافة إلى نهج التبسيط والارتقاء، الذي يحاول تخطى معضلة تعقيد الحياة، في وقت كانت فيه دراسة الطبيعة طريقا إلى معرفة الله. فكان مشاهير علماء التاريخ الطبيعي يلتمسون التقوى من خلال ذلك، مثل جلبرت وايت Gilbert White الذي ألّف كتابا عن اكتساب المعرفة بالملاحظة عنوانه (التاريخ الطبيعي لسلبورن The Natural History of Selborne)، والباحث الموسوعي ويليام هيول William Whewell الذي ساهم بأطروحته (دراسة علم الفلك والفيزياء العامة في ضوء اللاهوت الطبيعي) ضمن (أطروحات بريدچووتر Bridgewater Treatises)، والعالم الشهير ويليام بالي الذي ألف الكتاب الشهير (اللاهوت الطبيعي Natural Theology)، والذي أسس فيه لفكرة التصميم، وطرح حجته الشهيرة عن صانع الساعات Watchmaker analogy. وملخصها أنه إن كانت الصخرة الملقاة لا تلفت الانتباه لوجود التعقيد، لكن الساعة الملقاة على الأرض تلفت الانتباه وتشير إلى وجود صانع لها، والشيء نفسه ينطبق على الكائنات الحية.
في أواخر القرن التاسع عشر أتت الداروينية حاملة في كنفها نزعة مادية عاتية، كمخرج من سطوة الفكر الديني الأنجليكاني الذي تسيد المشهد البيولوجي آنذاك، فأنكرت وجود الغاية الكونية، واستبدلتها بمفهوم الانتخاب الطبيعي لقيادة التنوع الأحيائي، من خلال مبدأ الصراع من أجل البقاء، بالإضافة إلى نهج التبسيط والارتقاء، الذي يحاول تخطى معضلة تعقيد الحياة، في وقت كانت فيه دراسة الطبيعة طريقا إلى معرفة الله. فكان مشاهير علماء التاريخ الطبيعي يلتمسون التقوى من خلال ذلك، مثل جلبرت وايت Gilbert White الذي ألّف كتابا عن اكتساب المعرفة بالملاحظة عنوانه (التاريخ الطبيعي لسلبورن The Natural History of Selborne)، والباحث الموسوعي ويليام هيول William Whewell الذي ساهم بأطروحته (دراسة علم الفلك والفيزياء العامة في ضوء اللاهوت الطبيعي) ضمن (أطروحات بريدچووتر Bridgewater Treatises)، والعالم الشهير ويليام بالي الذي ألف الكتاب الشهير (اللاهوت الطبيعي Natural Theology)، والذي أسس فيه لفكرة التصميم، وطرح حجته الشهيرة عن صانع الساعات Watchmaker analogy. وملخصها أنه إن كانت الصخرة الملقاة لا تلفت الانتباه لوجود التعقيد، لكن الساعة الملقاة على الأرض تلفت الانتباه وتشير إلى وجود صانع لها، والشيء نفسه ينطبق على الكائنات الحية.
بيد أنه من المفارقات المتعلقة بهذا الشأن أن أحد أهم أسباب سقوط الحياتية هو ما ثبت صحته لاحقا مع سيرورة التقدم العلمي، فبالرغم من فشل الحياتيون في حل لغز تلك المادة المميزة للحياة، والتي أطلقوا عليها آنذاك البروتوبلازم، فإن المشتغلين بالبيولوجيا اليوم يعلمون أن الحياتية كانت على حق في احتجاجها بتميز الكائن الحي عن الجماد بميزات مذهلة، تم الكشف عنها تباعا مع تقدم العلم، وكان أهمها البرنامج المعلوماتي الجيني (الدي إن إيه DNA).
- ما هو النظام الحيوي؟
في عام 1931 أعلن عالم الوراثة والأحياء التطورية چون هالدِن J. S. Haldane أن علماء الأحياء لم يجدوا بدا من التخلي عن الحياتية بسبب كونها فكرة ميتافيزيقية في التوصيف المادي. لكنه في الوقت ذاته كان يقول إن أي تحليل آلي بحت لا يمكنه تفسير هذا التناسق المذهل للحياة. بذلك فإنه رغم انحسار فكرة الحياتية عن قيادة الوجهة البيولوجية، لم تتسيد فكرة الآلية الميكانزمية التسطيحية على المشهد، بقدر ما مهد لظهور مدرسة أخرى تسلمت الراية من سابقتها، ونحت وجهتها المادية بإتباع النهج الفيزيوكيمائى لتفسير النظام الحيوي، وسميت بالعضوانية. بالرغم من أن هذا النهج قد تأهل لتوصيف الكثير من العمليات الحيوية على مستوى الجزيئات، فإنه يفتقد صلاحيته تماما في تفسير علة ذلك التكامل التكويني المذهل للكائن الحي، وكيفية وسبب حدوثه. وكلما ارتفع المستوى الذي يجري فيه الاختبار على طريق تكامل تكوين الكائن الحي، ارتفع معدل الفشل وخابت الآمال المرجوة. فبعد النظر في المجهر، تبين للعلماء أن حياة الكائنات المتنوعة، من بشر وحيوانات ونباتات، هي محصلة لتعاون مئات المليارات من الخلايا المنفردة الدقيقة غير المرئية، التي تخصصت في وظائفها تخصصا عاليا، لدرجة أن أيًّا منها لم تعد قادرة على الحياة منفردة، من ثم أصبحت مهمة العلماء هي فهم وظائف الخلايا المنفردة وطريقة تعاونها؛ لأن المجال المرئي من العالم لم يقدم تفسيرا للحياة. وبدا لهم آنذاك، أن من يستطيع أن يعرف لماذا تمكنت هذه الملايين من الخلايا –التي توالدت جمعيها لدى كل حي من خلية بويضة ملقحة– من أن تتطور تطورا غائيًّا إلى العديد من الأنواع المختلفة من الخلايا عالية التخصص الوظيفي، فإنه بذلك قد ملك سر الحياة.
بيد أن هذا السر يأبى إلا أن يزداد غموضا فوق غموضه، فلا زالت مسألة التنوع الخلوي بدون حل حتى يومنا هذا، وما زاد الأمور تعقيدا، هو اكتشاف العلماء طبقة أعلى من التعقيد تحت المستوى الخلوي بداخل عمق الخلية ذاتها، وكان هذا المسار خطوة أولى فيما يسمى مجال البيولوجيا الجزيئية، التي أعطتنا الكثير من التساؤلات والألغاز المركبة، بدلا من أن تمنحنا الإجابات.
ظل التساؤل: كيف تنجز هذه الخلية مهامها؟ وما هي العوامل التي تنظم وظائفها المتعددة في كلٍّ واحدٍ منسجم؟
وصف العلماء الكائنات الحية كنظم غاية في التعقيد، على مستويات وطبقات متعددة؛ حيث تعتمد الخصائص المميزة لها على (تنظيم الكيان) أكثر من اعتمادها على (تركيب الكائن)، فارتباط الكل بأجزائه في عالم الحياة لا يقتصر على التكامل الكمي بينهما، بل يشمل أيضا ما ينتج عن ذلك من سيطرة الكل على أجزائه، وفهم الكائنات المتعضية الحية ينبغي أن يتم من منظور كلي. فالترابط بين أجزاء أي كائن حي متعض ينطوي على نوع من التفاعل النشط بينها، فهذه الجزيئات يمكن اعتبارها كليات صغرى، كما هو الحال في الخلايا التي يتألف منها جسم أي كائن حي. يشرح أليكس نوفيكوف Alex Novikoff: "إنّ الكلَّ والجزء كليهما كيانٌ مادي، والتكامل ينتج مما يتم بين الأجزاء من تفاعل مرتب على خصائصها". فالكلية لا تنظر إلى الوحدات الفيزيوكيميائية التي يتكون منها الكائن الحي باعتبارها أجزاء في آلة يمكن فكها ووصفها من دون اعتبار للجهاز الذي انتُزِعت منه، وهو ما لخصه بيولوجيون آخرون في عبارة موجزة: الكل، شيء يختلف عن مجموع أجزائه.
من هنا فإن وصف الأجزاء المعزولة لا يمكن أن ينقل صورة عن خصائص الجهاز الحيوي ككل، ولا يمكنه أن يفسر وجوده، فالذي يتحكم في الجهاز كله هو ما يربط بين هذه الأجزاء من نظام يطلق عليه اسم المتعضي Organization.
وتكامل الأجزاء قائم على كل مستوى من مستويات التكوين؛ ففي الكائن المتعضي الواحد يتم التكامل في الخلايا، ثم بين تلك الخلايا، فالأنسجة، فالأعضاء، فالأجهزة العضوية التي بتكاملها يكتمل كيان الفرد.
كيف يعمل هذا النظام؟ وما هو سر وجوده؟ وهل نجحت الداروينية –حاملةُ راية التفسير المادي– في شرحه؟ أم أن هناك نظريات أخرى بديلة؟
هذا ما سنحاول استكشافه من خلال التسلسل التالي.(2)(3)
- التعقيد الحيوي في مواجهة الداروينية
عندما رأى داروين التشابه في العضلات وبنية الجسم عبر العديد من الأنواع، لم يكن لديه المعرفة الكافية بهذه التعقيدات الهائلة الكامنة داخل تلك الأجهزة في ذلك الوقت المبكر من تاريخ العلم، لكنه أدرك بالرغم من ذلك حجم الإشكالية التي تواجه فرضيته؛ بنية الأعضاء الحيوية المعقدة بداخل كيانات الأحياء، والتي أطلق عليها أجهزة "مفرطة الإتقان والتعقيد Extreme Perfection and Complication".
أمام روعة هذه التصاميم الحيوية يقف داروين عاجزا، ويكتب عن تركيب العين في كتابه (أصل الأنواع):
"إن الافتراض بأن العين، بكل ما فيها من قدرات فذة لتعديل التركيز وفق مسافات متباينة، والسماح بكميات مختلفة من الضوء، وتصحيح الانحراف الكروي واللوني، قد صاغها الانتخاب الطبيعي، هو على ما يبدو افتراض سخيف غاية السخف، وأنا أعترف بذلك".(4)
 منذ الوهلة الأولى، أعلنت الأعضاء الحيوية مفرطة الإتقان عن تحدٍ واضح لفرضية التطور التدريجي البطيء التي تبنتها الداروينية كتفسير مادي طبيعي لحدوث الحياة بمعزل عن التصميم. تفترض الداروينية أن هذه العضيات الحيوية تمر أثناء رحلة تطورها المزعومة بسلسلة من المراحل الوسيطة الطفيفة والمتتالية، يقوم خلالها الانتخاب الطبيعي بصياغة تكيفها تدريجيّا، بالحفاظ على تغيرات المرحلة المفيدة والوظيفية، وتدمير ما هو غير صالح أو أقل تكيفا. وهنا تكمن المعضلة المحورية، فهذه الأعضاء لا تستطيع أن تقوم بوظيفتها إلا بوجودها مكتملة، والكيانات الوسيطة المتتالية التي من المفترض أن يمر بها العضو أثناء رحلة تطوره ليس لها أي معنى وظيفي إلا بوصفها أجزاء من المنتج النهائي، لذلك فإن كافة المراحل الوسيطة ليس لها أي ميزة انتقائية، وينعدم الدور المخول للانتخاب الطبيعي في الحفاظ عليها وتثبيتها، لأنها لا تحمل له أيّة قيمة تكيفية أثناء تطور العضو، بل هي مجرد أعضاء مشوهة ناقصة تمثل عبئ يجب التخلص منه.
منذ الوهلة الأولى، أعلنت الأعضاء الحيوية مفرطة الإتقان عن تحدٍ واضح لفرضية التطور التدريجي البطيء التي تبنتها الداروينية كتفسير مادي طبيعي لحدوث الحياة بمعزل عن التصميم. تفترض الداروينية أن هذه العضيات الحيوية تمر أثناء رحلة تطورها المزعومة بسلسلة من المراحل الوسيطة الطفيفة والمتتالية، يقوم خلالها الانتخاب الطبيعي بصياغة تكيفها تدريجيّا، بالحفاظ على تغيرات المرحلة المفيدة والوظيفية، وتدمير ما هو غير صالح أو أقل تكيفا. وهنا تكمن المعضلة المحورية، فهذه الأعضاء لا تستطيع أن تقوم بوظيفتها إلا بوجودها مكتملة، والكيانات الوسيطة المتتالية التي من المفترض أن يمر بها العضو أثناء رحلة تطوره ليس لها أي معنى وظيفي إلا بوصفها أجزاء من المنتج النهائي، لذلك فإن كافة المراحل الوسيطة ليس لها أي ميزة انتقائية، وينعدم الدور المخول للانتخاب الطبيعي في الحفاظ عليها وتثبيتها، لأنها لا تحمل له أيّة قيمة تكيفية أثناء تطور العضو، بل هي مجرد أعضاء مشوهة ناقصة تمثل عبئ يجب التخلص منه.
الانتخاب الطبيعي عملية لا غرض لها، عمياء عن رؤية المستقبل، ليس لها أهداف، ومعيارا التقييم الوحيدان لها هما النجاح في البقاء، والنجاح في التكاثر. وهذا ما يجب أن يتوافر في كل خطوة من خطوات التغيير في نشوء العضو الحيوي، لكن الطبيعة غير الاختزالية للعضيات الحيوية التي لا تقبل التدرج الوظيفي أو الإنقاص، تُفْشِل هذه العملية تماما، فهي إما تكون ككل أو أبدا لن تكون.
بالرغم من اعتراف داروين بحجم تلك الإشكالية، فقد تعاطا معها بمنهجية ملتوية، ونقل عبئ الإثبات لجهة المشككين؛ مطالبا إياهم بإثبات خطأ ادعائه الذي من المفترض أن يطالب هو بإثباته كأصل لقبول فرضيته، وذلك لإدراكه الراسخ بأنها جولة خاسرة، فقال في كتابه (أصل الأنواع):
"إذا كان من الممكن إثبات وجود أي عضو معقد لا يُرجَّح أنه قد تشكل عن طريق العديد من التعديلات المتعاقبة والطفيفة، فسوف تنهار نظريتي تماما".(1)
علينا أن ندرك أن داروين كان حذقا في التعامل مع العقبات التي تعترض فرضيته، واستعمل المواربات الذكية، على أمل أن تحل تلك العقبات آجلا، وفي المقابل سلم بالانهيار التام لفرضيته إذا لم تتمكن من تخطيها.
مرت عقود عديدة على طرح داروين، وهو ما يمثل زمنا طويلا جدا بمقياس مسار العلم الحديث، ويبقى التساؤل البَدَهيّ الذي يفرض نفسه حتى الآن؛ هل نجح تلاميذه في تخطي تلك العقبات؟
- العقبة الكبرى
مع التقدم العلمي الهائل في رصد تعقيدات حيوية مذهلة، كان من المفترض أن تزيد صعوبات تفسير تلك التعقيدات بالارتقاء والتدرج في هذا العصر، مقارنة بعصر داروين. إلا أننا –على النقيض من ذلك– نلاحظ أن داروين كان أكثر معقولية واتساقا من أتباعه في مواجهة مشكلات التعقيد. ترفع داروين عما نراه حاليا من سلوكيات متعمدة من أتباعه تمارس التدليس الانتقائي بتسطيح التناول حول تطور هذه الأعضاء. ويمكننا القول، أن من السذاجة قبول أن مقارنة بسيطة لسلسلة من الكائنات الحية الأقل تعقيدا بأخرى أكثر تعقيدا –وفق معرفتنا الحالية بكم هذا التعقيد– ستقنعنا بأن نوعا من التطور التدريجي على مدى ملايين السنين هو المسؤول عن تفسير التعقيد الحادث. وهذا السبيل بعينه هو ما يجادل عنه أنصار التطور منذ عهد داروين وحتى يومنا هذا، لتفسير وجود تلك النظم الحيوية المعقدة، دون تقديم أيّة آلية فعلية لسيرورة هذا التحول المفترض. فعلى سبيل المثال، تعرض لنا الداروينية محاكاة لتطور العين، وتشرح كيفية تحول بقعة حساسة للضوء –كنظام رؤية بدائي– تدريجيا إلى تجويف منخسف شديد التقعر، ثم نصف كرة مملوءة بمادة شفافة، ثم بعد ذلك تحولت إلى ما هي عليه الآن في العيون الأكثر تعقيدا، معتمدا في ذلك على صنع اصطفاف لنماذج من العيون المختلفة في الكائنات الحية اليوم، من الأبسط إلى الأعقد.
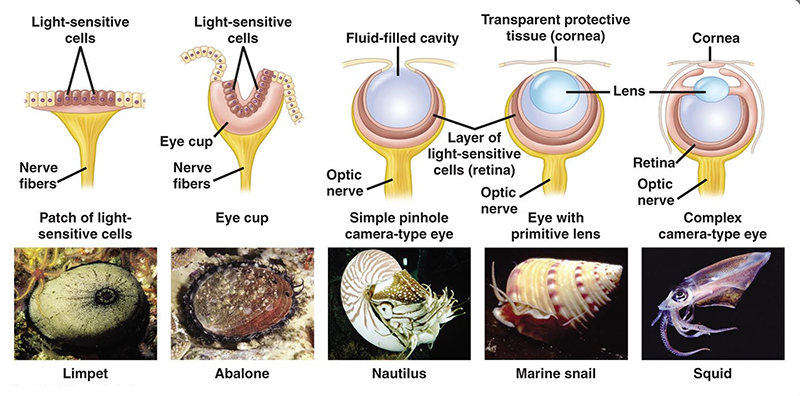
تكمن الإشكالية هنا في خطأ منهجي، متمثل في صحة الاستدلال على التطور بالاستناد على خصائص بعض العيون الموجودة في الوقت الحاضر، فلا يمكن بأي حال من الأحوال، تمثيل العيون التاريخية التي من المفترض أن تمثل الأسلاف، من خلال سرد سلسلة من العيون الموجودة حاليا، والتي تمتلكها أنواع حية عصرية. لأن التطور يتطلب أن كل العيون الحالية لديها نفس القدر من التاريخ التطوري، فهي كأبناء عمومة لا يمكن اعتبارها أسلافا لبعضها البعض.
إن هذا النهج يتطلب اعتراف الداروينية بأن هذه العيون الحديثة –البسيطة– في أول السلسة التطورية هي نظائر للعيون المفترضة للأجداد، ومماثلة لها، وهذا يضعهم في حرج حقيقي؛ لأنه بالتبعية يؤكد أن تلك العيون لم تتطور أو يطرأ عليها أي تغيير منذ أزمان سحيقة، وهو ما يناقض الفرضية التطورية برمتها حول سيرورة التطور الدؤوبة. داروين نفسه كان يدرك جيدا عدم صلاحية هذا الاصطفاف للاستدلال، وعلى الرغم من ذلك وقع هو نفسه في ذلك الخطأ، وتفشى في نهج أتباعه.(5)
حين نحاول التعاطي مع حجة التطور السابقة حول اصطفاف تطور العين كمثال، فإننا بمجرد الخوض في أولى خطوات اختبارها، نصطدم مباشرة بالكثير من التناقضات، ونرصد خرقا جسيما لما تضعه الداروينية لنفسها من آليات وإلزامات وقواعد داخل إطار الاستدلال العام على التطور. وهذا ليس مستغربا، لكن على الرغم من ذلك لا ييأس أتباعها من البحث عن مبررات لتخطيها، وتقع أغلبها تحت إطار الحجج الدائرية، أو المسكنات الموضعية للإشكالية، أو ما يمكن تسميته بمبررات الاستهلاك المحلي، فحين ننظر في محاذاة عيون الأنواع الحية من البسيط إلى الأكثر تعقيدا لتبرير تطور العين، نجد أنها تناقض شجرة التطور الفيلوجينية التي تم رسمها.
على سبيل المثال: تخبر الداروينية أن أسلاف الرخويات قد انشقت عن الفقاريات خلال عصور سحيقة من تاريخ الحياة، في عصر ما قبل الكامبري Precambrian، مما يجعل البشر أكثر ارتباطا بنجم البحر وديدان الأرض، عن ارتباطهم بالأخطبوط (من الرأسقدميات)، بناء على ذلك فمن المفترض أن نرى العيون البسيطة البدائية في السلسلة متواجدة لدى الأخطبوط، لكن من الحقائق المثيرة للدهشة أن الأخطبوطات بالرغم من انحدارها في أسفل سلسلة التطور، إلا أنها تمتلك ذلك النوع من نظام الرؤية شديد التعقيد، والمعروف بعيون الكاميرا الذي يميزنا نحن البشر، والعجيب هنا أن التشابه بين عيون الإنسان والأخطبوط –رغم المسافة التطورية الشاسعة بين كلا النوعين– يبدو متطابقًا في التراكيب على نحو مذهل.
التطابق البنيوي بين عين الأخطبوط وعين الإنسان لغز يتحدى العشوائية
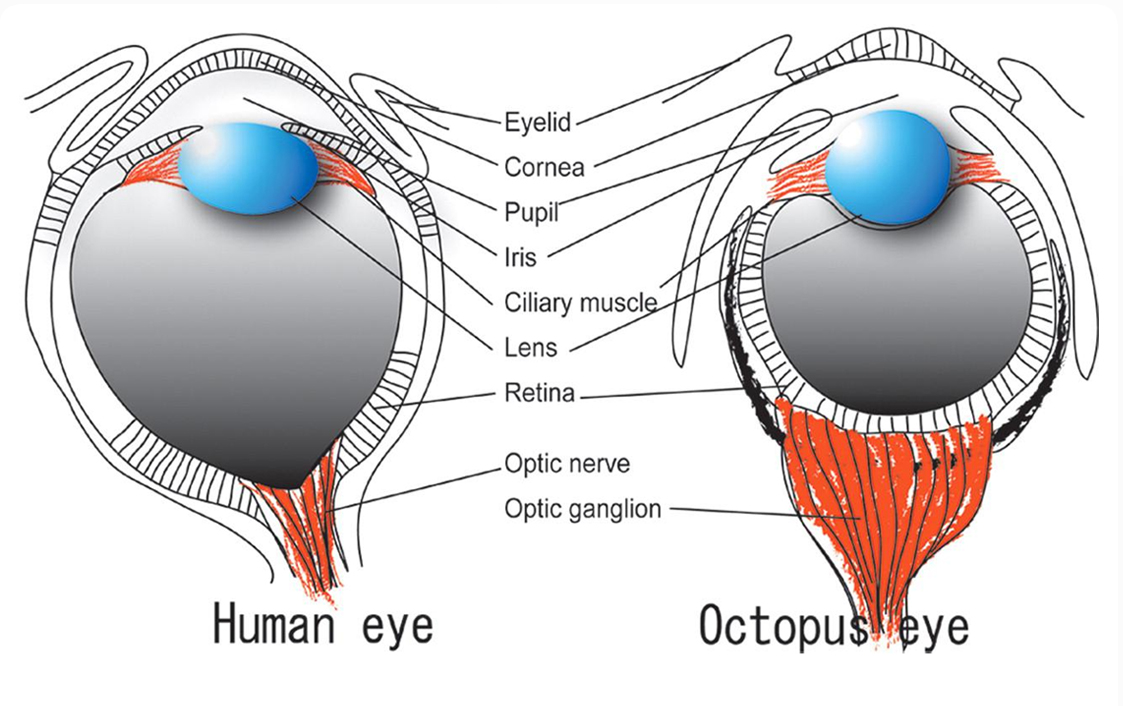
genome.cshlp.org/content/14/8/1555/F1.expansion.html
هذه القفزة الهائلة في تشكل العين عبر سلسلة المحاذاة، لا يمكن تفسيرها بالانحدار من أي سلف مشترك، وتقف مباشرة في وجه السيناريو التطوري الذي يستدل بتدرج نماذج العيون الحالية كتمثيل لأسلافها، وكالعادة، لا يمكن لأنصار التطور تحمل تبعات فرضياتهم، فنجدهم يتملصون منها بالمجادلة حول كون كل من الأخطبوط والإنسان أبناء عمومة من سلف قديم جدا، عاش في عصور ما قبل الكامبري، ولا يمثل أحدهما سلفا للآخر، وتلك البنية المعقدة والمتطابقة في نظام الرؤية بين كلا النوعين قد تطورت بمعزل عن بعضها البعض بطريق تطوري تقاربي Convergent Evolution.(6)
هذا الرد هو ما يعنينا الآن من طرح المثال السابق، وعليه نتساءل: إن كانت الكائنات الحية لا يمكنها أن تمثل الأسلاف ولا تعبر عنها، فلماذا يحتج أنصار التطور بمصفوفات كائنات حية حالية لتبرير تسلسل تطور العين؟!
المثال السابق ليس استثناءً، فقد رصدت التناقضات على نطاق كبير، حيث لاحظ العلماء أن العيون في ثلاثة فئات رئيسية من الأنواع (الفقاريات والمفصليات والرخويات) تنشأ من أنسجة مختلفة، ومنه افترض أنصار التطور أن لدى هذه الفئات الثلاثة تاريخا تطوريا منفصلا، والعديد من أوجه التشابه بينها نتجت أيضا بسبب التطور التقاربي.(7)
مثل هذه التناقضات الشديدة في المسار التاريخي المقترح لتطور العين، دعت أنصار التطور إلى افتراض أنها قد تطورت ونشأت أكثر من أربعين مرة بشكل مستقل، وهو ما يفسد سيناريو الاصطفاف، ويؤكد عدم جدواه في الاستدلال.(8)(9)
بالإضافة إلى تناقض هذا النموذج مع السجلات الجيولوجية التي لا تدعم هذا التدرج المزعوم، وتستعرض ظهورا فجائيًّا للعيون المعقدة، بكامل تراكيبها المعروفة اليوم، منذ أكثر من 540 مليون سنة فيما عرف بالعصر الكامبريCambrian، ويشير إلى ثباتها منذ وجودها للوهلة الأولى، ولا زال أصلها التطوري يمثل لغزا حقيقيا.(10)(11)(12)
بتخطي هذا النهج المعيب، نتحول إلى محور تناولنا الذي يتعلق بتلك الإشكالية الكبرى التي تتعامى عنها الداروينية، وهي آلية هذا التحول المطلوب لإنتاج ذلك الجهاز المعقد العين.
فحدوث كل طفرة متتابعة في الحمض النووي داخل مقلة العين يتطلب تحولات جذرية ومتزامنة وشديدة التعقيد، والترابط داخل بنية العظام والأعصاب، ووظيفة الدماغ، ويجب سلوك مئات من المسارات التطورية في ذات الوقت عن طريق طفرات فاعلة في جميع الجوانب المتعلقة بالرؤية، مثل هذه التغييرات تتطلب أكثر بكثير مما يمكن توقعه من الطفرات العشوائية والانتخاب الطبيعي.(13)
ولتقريب مدى صعوبة تلك العملية باتخاذ مقياس واحد فقط، هو عدد الجينات المسئولة عن إنتاج العين، نجد أنه قد تم التعرف حتى الآن على 501 جين مرتبط بالعين في الحشرة البدائية (ذبابة الفاكهة)، أي ما يعادل 3.5٪ من حجم جينومها بأكمله.
وفي الكائنات الأكثر تعقيدا مثل الفقاريات، نجد أن أكثر من 7500 جين يتداخل في تركيب وتنظيم شبكية العين، أي حوالي 30٪ من الجينات البشرية قاطبة، لذلك فحدوث طفرات متزامنة على هذا العدد الهائل من المسارات والجينات هو فرضية مريعة، تتخطى حدود المنطق، وتتزايد مع مستويات التكامل الأرقى بين أجزاء تشكل العين. حتى على المستويات التكوينية الأقل، تدرك الداروينية تلك المشاكل الخطيرة في وضع تفسير لتطور كل جزء من نظام الرؤية، بما في ذلك العدسة، ومقلة العين، وشبكية العين، والنظام البصري بأكمله، وفصوص القذالي في الدماغ.(14)
مثل هذا التكامل والتنظيم دفع عالم الفسيولوجيا سكوت تيرنر J. Scott Turner لتسمية عملية الرؤية بالمعجزة، ويقول إن معجزة الرؤية الحقيقية تكمن في تلك العملية الحسابية التي يمكن أن تنتجها.(15)
كل هذه الأنظمة المختلفة يجب أن تعمل معا كوحدة متكاملة لتحقيق الرؤية. في بحث خاص يدرس أعضاء أقل تعقيدا في تشريح أسد النمل، يتساءل بيريه جراسي Pierre P. Grassé حول إمكانية إنتاج مثل هذه النظم المعقدة بواسطة الانتخاب الطبيعي الذي يعمل على الطفرات العشوائية، وحول فرص حدوث مثل هذه الطفرات المتزامنة التي يمكنها أن تفعل ذلك، وجدوى هذه الطفرات في إنتاج الهياكل التي تلائم بعضها البعض بدقة.(16)
من المؤكد أن مثل تساؤل جراسي سيلاقي طريقا مسدودا، فحتى أبسط نظم الرؤية وأكثرها بدائية (البقع الحساسة للضوء) –التي استغلت كحلقات أولية في سلسلة التطور– تتطلب وجود مجموعة كبيرة ومعقدة من النظم الإنزيمية في المكان والوقت المناسب لكي تعمل، وهي بحد ذاتها تعقيد لا يصدق، ونظام لا يمكن اختزاله، فعندما اتخذ دوكينز وغيره من أنصار التطور البقعة الحساسة للضوء كبداية للصعود نحو سفح الجبل، وقعوا في مغالطة منطقية بسبب تعقيدها الهائل، وبذلك فرحلته نحو القمة محكوم عليها بالفشل؛ لأنه ابتدأ من قمة أصلا، وهوى مباشرة نحو الهاوية.
 في ورقة نشرت مؤخرا من المركز الطبي بجامعة بايلور بعنوان (تشريح الداروينية)، يشير الدكتور جوزيف كونJoseph A. Kuhn إلى أن الكثير من الأطباء خلال دراستهم للتعقيد الهائل للجسم البشري يمكنهم تقبل حدوث انتقاء لبعض الطفرات التي تعمل على مقاومة الملاريا، وخصائص الجلد، والعديد من التغييرات الطفيفة الأخرى التي لا يمكنها تحويل النوع. لكن مثل هذه الطفرات لا تقدم أي تفسير حقيقي حول منشأ وتشكيل الأجهزة والنظم المعقدة، فجميع عناصر النظم الحيوية تقريبا يجب أن تكون موجودة في وقت واحد بدلا من أن تتطور تدريجيا، فيما أسماه نظام (كل شيء أو لا شيءAll Or Nothing ).
في ورقة نشرت مؤخرا من المركز الطبي بجامعة بايلور بعنوان (تشريح الداروينية)، يشير الدكتور جوزيف كونJoseph A. Kuhn إلى أن الكثير من الأطباء خلال دراستهم للتعقيد الهائل للجسم البشري يمكنهم تقبل حدوث انتقاء لبعض الطفرات التي تعمل على مقاومة الملاريا، وخصائص الجلد، والعديد من التغييرات الطفيفة الأخرى التي لا يمكنها تحويل النوع. لكن مثل هذه الطفرات لا تقدم أي تفسير حقيقي حول منشأ وتشكيل الأجهزة والنظم المعقدة، فجميع عناصر النظم الحيوية تقريبا يجب أن تكون موجودة في وقت واحد بدلا من أن تتطور تدريجيا، فيما أسماه نظام (كل شيء أو لا شيءAll Or Nothing ).
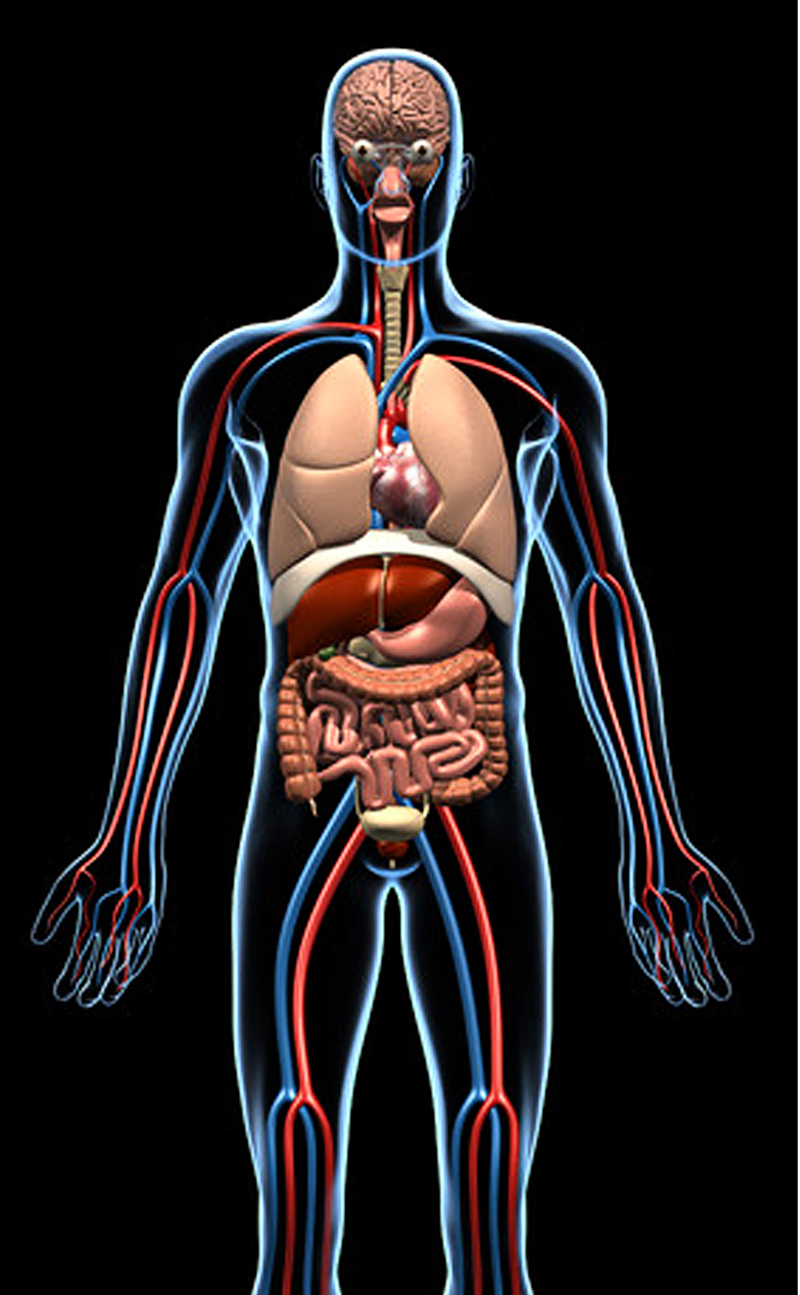
أمثلة عديدة من داخل الجسم البشري للأنظمة المعقدة المتخصصة غير القابلة للاختزال، التي لا يمكن تشكلها من قبل الطفرات المتتابعة، حيث يتوجب على جميع المكونات أن تكون موجودة لتعمل تلك الأنظمة بشكل صحيح، وتشمل هذه النظم المعقدة (الرؤية، التوازن، الجهاز التنفسي، الجهاز الدوري، الجهاز المناعي، الجهاز الهضمي، الجلد، نظام الغدد الصماء، الذوق)، وغيرها من الأمثلة على المستويات البيوكيمائية والتشريحية ووظائف الأعضاء.
يشير كوِن إلى أن الداروينية لا يوجد لديها تفسيرات فعلية لأصل النظام المعقد الذي لا يمكن اختزاله، ناهيك عن شبكة مترابطة من الأنظمة غير القابلة للاختزال، التي تشكل جسم الإنسان ككل. بالتالي فجسم الإنسان يمثل نظامًا معقدًا لا يمكن اختزاله على النطاق الخلوي والأجهزة والنظم.(17)
- كل شيء.. أو لا شيء (التعقيد غير القابل للاختزال)
 يستعرض مايكل بيهي أستاذ الكيمياء الحيوية في جامعة ليهاي بــبنسلفانيا، وأحد منظري التصميم الذكي، من خلال كتابه الشهير (صندوق داروين الأسود: تحدي الكيمياء الحيوية للتطور) طبقة أخرى من النظم غير القابلة للاختزال على المستوى البيوكيمائي الجزيئي في الحياة المجهرية الدقيقة.
يستعرض مايكل بيهي أستاذ الكيمياء الحيوية في جامعة ليهاي بــبنسلفانيا، وأحد منظري التصميم الذكي، من خلال كتابه الشهير (صندوق داروين الأسود: تحدي الكيمياء الحيوية للتطور) طبقة أخرى من النظم غير القابلة للاختزال على المستوى البيوكيمائي الجزيئي في الحياة المجهرية الدقيقة.
ويعرّف بيهي النظام غير القابل للاختزال Irreducible complex بأنه نظام مركب من العديد من الأجزاء التي تتفاعل بتناسق شديد لإنتاج الوظيفة المخولة بالنظام، ويتبع ذلك أن إزالة جزء واحد من أجزائه يعطل هذا النظام ويوقفه عن العمل.
مما يعني أنه قد تم تصميم جميع أجزائه من البداية، بذلك لا يمكن أن يتكون نتاجا لطفرات طفيفة متدرجة يتم انتخابها، فالتطور لا يمكنه بناء عمليات بيوكيميائية معقدة خطوة بخطوة، لأن تلك الخطوات لا يمكنها توفير أيّة ميزة لحاملها، هذا يعني أن الانتخاب الطبيعي لن ينتقي هذا النمو التطوري، ولن يسمح لنظام غير كامل وغير فعال بالانتشار من جيل لآخر، فهو لا يثبت سوى التغيرات الوظيفية، وهو ما تفتقده تلك النظم، التي لا تعرف وظيفتها إلا بوجودها مكتملة.
يستخدم بيهي مصيدة الفئران كمثال تقريبي لشرح فكرته حول التعقيد غير القابل للاختزال، فهي لا يمكن أن تعمل إذا فقدت أيًّا من أجزائها المكونة من (القاعدة، الزنبرك، الماسك، المطرقة)، ولا يكفي وجود كل الأجزاء في الوقت والمكان المناسبين، بل يجب أن تكون متناسقة مع بعضها بدقة، حتى يمكنها القيام بوظيفتها، مما يجعل فرضية الفرص العشوائية محض جنون.
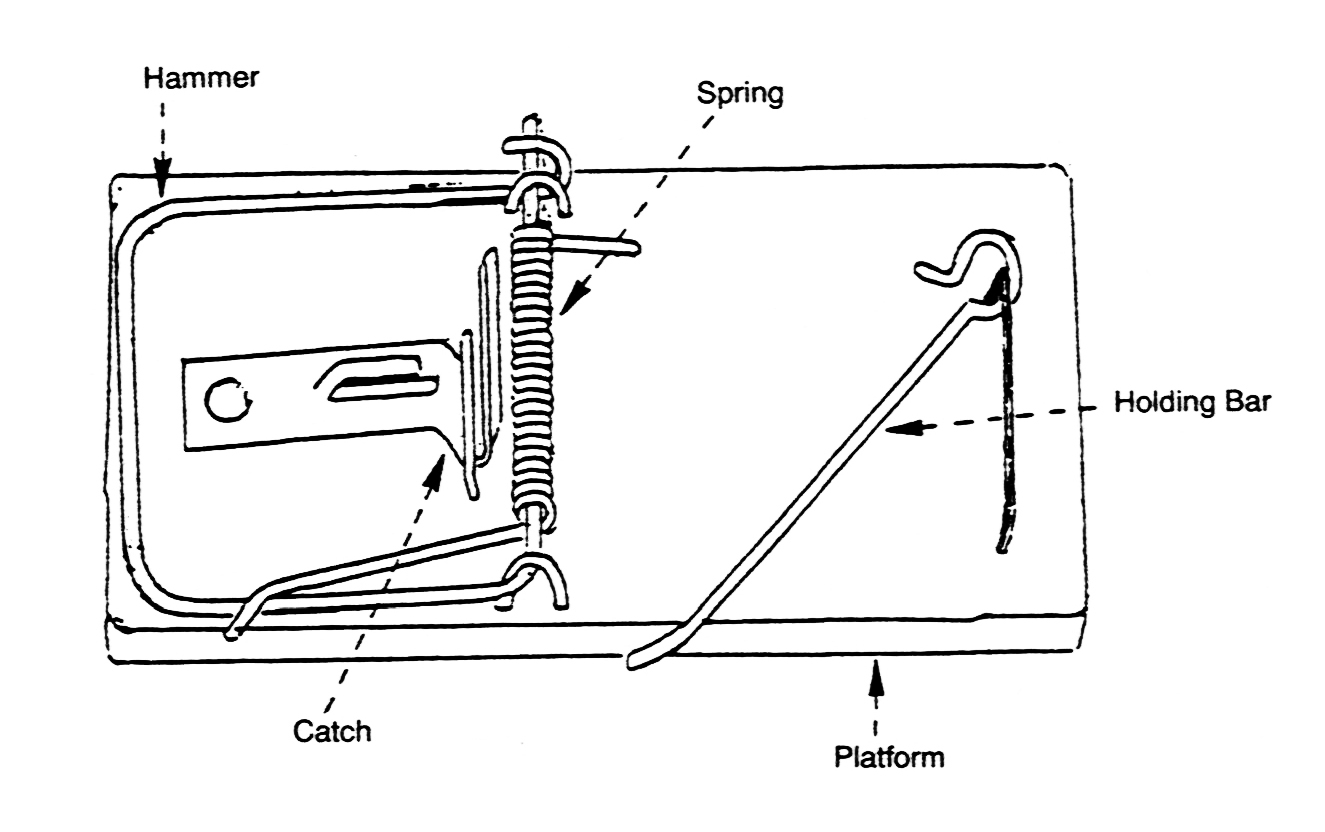
ينتقل بيهي لشرح العديد من تلك النظم البيوكيميائية، منها آلية استشعار الضوء في نظام الرؤية، ونظام تخثر الدم البشري، والسوط البكتيري الذي تستخدمه الكثير من البكتيريا للحركة.
فالعين تحمل طبقة أخرى من التعقيد على المستوى البيوكيميائى الدقيق، وعندما يضرب الضوء الخلايا الحساسة في العين، تحدث سلسلة من التفاعلات الجزيئية المتعاقبة المذهلة تماما، التي يمكن تشبيهها بانهيار قطع الدومينو المتراصة حين نسقط أول قطعة، لتؤدي في النهاية إلى نقل النبض العصبي إلى الدماغ.
وإذا ما فُقِدَ أيُّ جزيء في هذه السلسلة من قطع الدومينو أو كان معيبا لن يمكن بث النبض العصبي، مما يعنى ببساطة انعدام الرؤية والعمى التام، لذلك فإن هذا النظام لا يمكنه العمل على الإطلاق ما لم تكن كل أجزائه حاضرة في ذات الوقت.
في مثاله الثاني شلال تجلط الدم في الإنسان، والمسئول عنه أكثر من عشرة جزيئات بروتين شديدة التناسق، يجب أن تتفاعل مع بعضها البعض بشكل متسلسل لإنتاج الجلطة في الوقت والمكان المناسبين لإيقاف نزيف محتمل، وفقد أحد هذه الجزيئات يعنى فشل نظام التخثر تماما –كما في حالة مرض الهيموفيليا–.
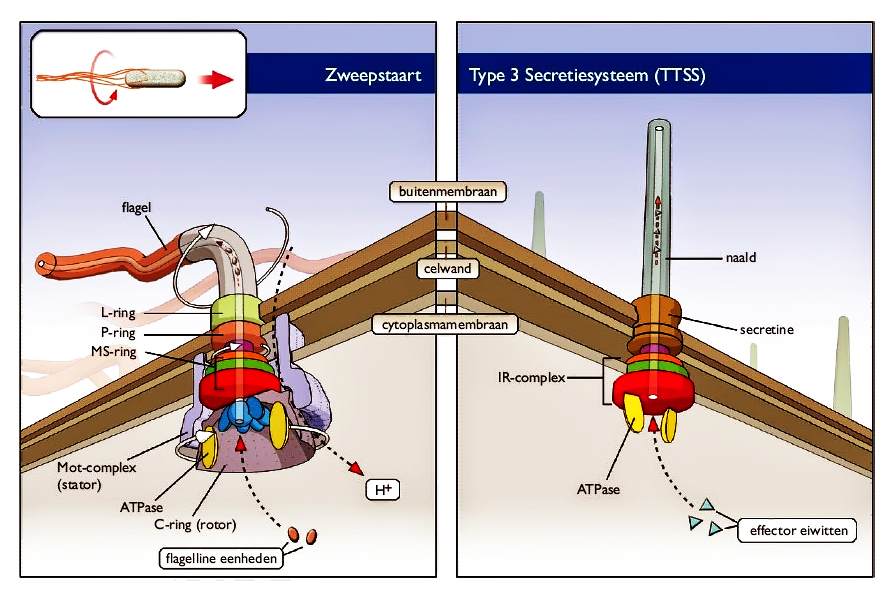
المثال الثالث هو السوط الباكتيري Bacterial Flagellum، الذي يبدو مثل خيوط خارجية طويلة تساعد على دفع البكتيريا، وتمكنها من السباحة والحركة خلال السائل المحيط بها. يتحرك السوط بآلية ميكانيكية تماما، فعند قاعدة كل سوط محرك دوار يحركها آلاف المرات في الدقيقة الواحدة.
يتكون السوط البكتيري من الخيط Filament الذي يرتبط مع الجسم القاعدي Basal body بواسطة قطعة وسطية تسمى الخطاف Hook.
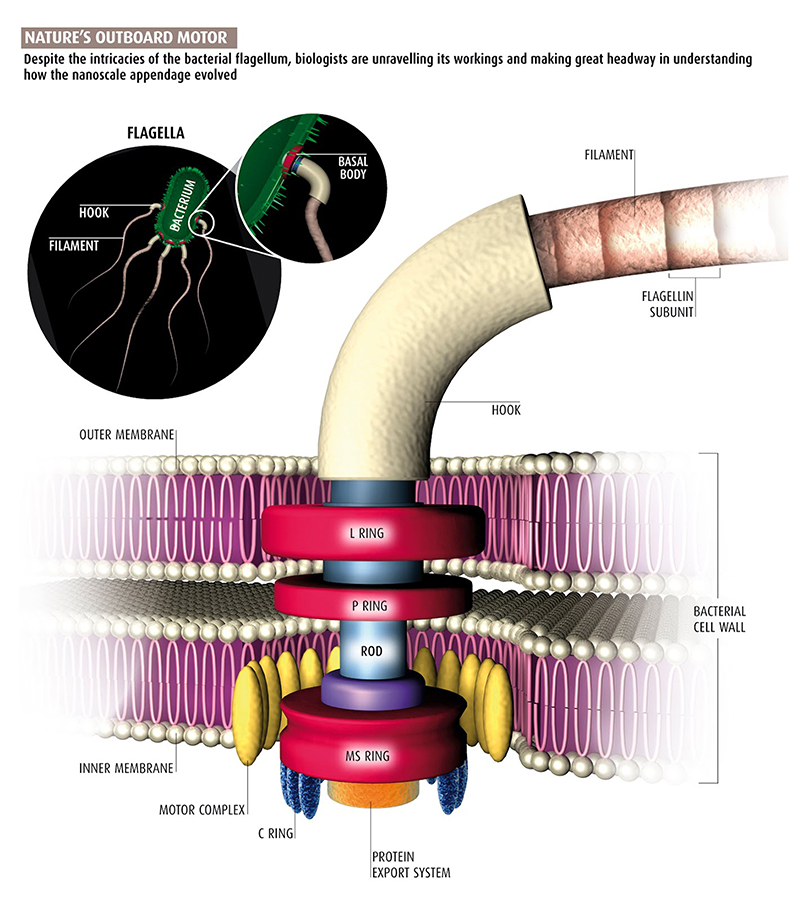
يحاكي السوط البكتيري أنظمة الحركة الميكانيكية بوسائل النقل المائية بآلية الدفع اللولبي، بواسطة محركات دوارة لا يمكنها العمل مع فقد أحد أجزائها، ومثل هذا النظام لا يقبل التدرج في التكوين، فالتطور بطريق الانتخاب الطبيعي والطفرة يجب فيه المضي الحثيث نحو تحسين وظيفية واحدة في كل خطوة، فكيف يمكنه بناء جهاز لا يمكن اختزال خطواته مثل المحرك الدوار الذي لا يمكن أن يعمل على الإطلاق إلا إذا كانت جميع أجزائه في مكانها الصحيح؟!(18)
- حجج الداروينية المضادة
في عام 2005، أقامت منظمة الدفاع عن حريات المواطنين الأمريكية American Civil Liberties Union بالاتفاق مع إحدى عشرة أسرة ممن يتعلم أبناؤهم في مدارس منطقة دوڤر التابعة لولاية بنسلفانيا، دعوى قضائية ضد المجلس التعليمي للمنطقة.(19)
كان السبب وراء ذلك هو أن مجلس إدارة مدرسة منطقة دوڤر قد قرر دعوة الطلاب من خلال بيان مقتضب يتلى عليهم لدقيقة، يعرفهم على أن هناك نظرية أخرى بجوار نظرية التطور وهي نظرية التصميم الذكي. وكان الأساس الذي بنيت عليه الدعوى هو أن التصميم الذكي ينطلق من أسس دينية، وبالتالي فتدريسه في حصص العلوم يخالف التعديل الأول من الدستور الأمريكي، والذي نص على أن الكونجرس لن يصدر قوانين بناء على أية أسس دينية.
كينيث ميلرKenneth Miller هو أحد الشهود الخبراء، ممن استعان بهم الادعاء في القضية، وظهر مايكل بيهي Michael Behe الشاهد الخبير للدفاع، أثناء استجواب ودّي طرحه عليه الادعاء، أكد ميلر أن نظرية التصميم الذكي "ليست قابلة للاختبار"، لذلك فهي ليست من العلم في شيء، لكن في وقت لاحق أثناء شهادته ناقض ميلر ادعائه هذا، وأثبت أنها نظرية قابلة للاختبار، حين جادل أن العلم قد اختبر حجة النظام غير المختزل وأثبت زيفها.
بالتغاضي عن تناقض ميلر الذي لم ينتبه إليه القاضي أو المحلفين في حينه، لنرى كيف أثبت زيف حجة بيهي حول التعقيد الذي لا يمكن اختزاله: يمكن للمطلع بسهولة أن يدرك أن ميلر ارتكز خلال شهادته وفي كتاباته العديدة على تشويه الحجج وطرح تعريف مضلل للنظام غير القابل للاختزال، ومن ثم يقوم بدحض هذه الفكرة المضللة والمشوهة التي صنعها بنفسه، مرتكبا بذلك حيلة منطقية يجيدها؛ تعرف بمغالطة (رجل القش).(20)
دلس ميلر في تعريف النظام غير القابل للاختزال، بادعائه أن حجيته تكمن في كون الأجزاء المكونة له لا تحمل أي معنى وظيفي خارجه، فبذلك يمكن تفنيده إذا ما تم العثور على وظيفة ثانوية لهذه الأجزاء في نظم أخرى، ويستخدم مثال بيهي (مصيدة الفئران) للبرهنة على ذلك بنزع أحد أجزائها واستخدامها كدبوس لرابطة العنق، ومن ذلك يدعي أن مفهوم التعقيد غير القابل للاختزال في السوط البكتيري قد تم دحضه، لأن ما يقارب ¼ البروتينات المستخدمة في وظيفة سوط البكتيريا تقوم بوظائف في نظام آخر في أنواع بكتيرية مختلفة، هو آلة حقن السم، وتسمى (نظام إفرازي النمط–III،أو T3SS)، مما يدل على أن السوط البكتيري من الممكن أن يتطور تدريجيًّا من نظام آلة الحقن الأقل تعقيدا.(21)
للتعريف بخدعة ميلر وتدليسه في اختبار النظام غير المختزل نضرب لكم هذا المثال: بما أننا قد وجدنا أن كلا من جهاز الكمبيوتر والموقد الكهربائي لديهما سلك الطاقة المسئول عن توصيل التيار الكهربائي، إذن نستنتج من ذلك أن جهاز الكمبيوتر لم يعد تعقيدا غير قابل للاختزال، وذلك لأن الكمبيوتر يتطلب عددا من قطع الغيار اللازمة من أجل وظائف ثانوية في أجهزة أخرى.
نعلم جيدا أن توافر بعض قطع جهاز الكمبيوتر الذي نستعمله الآن أو كل أجزائه في أي مكان آخر لا يغني عن تلك الخطوات الذكية التي يجب أن تكون مسئولة عن تجميع كافة الأجزاء في الوقت والمكان المناسبين، وفق مخطط إنشائي مسبق وتعليمات موجهة، والطريقة الصحيحة لتفسير تعقيده هي تتبع كافة خطوات تركيبه.
يضع كيسى لسكين Casey Luskin مقاربة تصويرية رائعة لمنطق ميلر السابق، في مقالة نقدية نشرها حول مجريات وأحداث محاكمة دوڤر، ممثلا في تفسير التعقيد غير القابل للاختزال من خلال شكل القوس.(22)
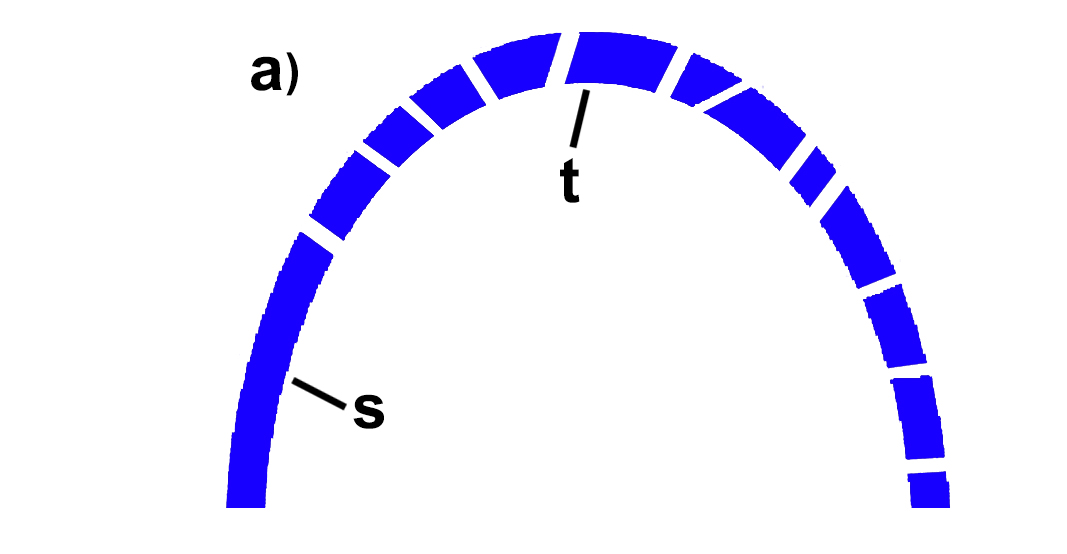
الشكل A: باعتبار القوس وظيفة معقدة لا يمكن اختزالها، ومقسم إلى العديد من القطع، بما في ذلك القطع T وS.

الشكل B: برفع القطعة T من القوس ينهار القوس، لكن تبقى القطعة S شاخصة ليس لها أيّة قيمة.
من خلال الرسم السابق نطرح سؤالا: هل فسر أو دحض وجود الجزء القاعدي (S) من القوس التعقيدُ غيرُ القابل للاختزال للقوس؟ بالطبع لا. وإذا قمنا بتمثيل القطعة (S) بقاعدة إبرة الحقن T3SS، فكونها مشتركه مع السوط لم يدحض التعقيد غير القابل للاختزال في تركيب السوط، وفشل تماما في تفسيره، الاختبار الحقيقي الوحيد الذي يمكنه دحض هذا النظام هو إظهار قدرة الطفرات العشوائية والانتخاب الطبيعي على بناء هذا النظام تدريجيا خطوة بخطوة، كما اشترط داروين، فبروتينات نظام حقن السم T3SS المشتركة مع السوط البكتيري لا تمثل أكثر من وظيفة المساهمة في تثبيت كلا منهما في غشاء الخلية، ولا تساهم في الوظيفة غير الاختزالية للسوط.
إذا ضربنا مثالا أكثر ملائمة، وقمنا بتشبيه السوط البكتيري بمحرك دفع خارجي لقارب، ونظام الحقن T3SS كرشاش مياه، فحتى نستطيع تثبيت كليهما على ظهر القارب –الذي يمثل بدوره جسم البكتيريا– يتوجب علينا استخدام قاعدة تثبيت (البروتينات القاعدية المشتركة)، ومنه يمكننا القول إنه من السذاجة استنتاج أن وجود جزء التثبيت القاعدي في رشاش الماء دليل على نفي التعقيد غير المختزل في تركيب المحرك الخارجي، مثل هذه الحجة –على حد تعبير ديمبيسكي– في أحسن الأحوال كمثل القول أن بإمكاننا السفر سيرا على الأقدام من لوس أنجلوس إلى طوكيو؛ لأننا اكتشفنا جزر هاواي.(23)
- ما هي متطلبات دحض نظام معقد غير اختزالي؟
لتتمكن الداروينية من وضع اختبار حقيقي لأصل آلة جزيئية متكاملة غير قابلة للاختزال وظيفيا، يتوجب عليها تتبع الخطوات والشروط الآتية، التي تم تلخيصها وفقا لمنيوجAngus Menuge:
أولا: توافر وإتاحة كل الأجزاء اللازمة لتشكيل النظام المطلوب.
ثانيا: تموقع الأجزاء بموقع البناء في الوقت الذي يتطلب وجودها فيه.
ثالثا: التنسيق والتوافق لتلك الأجزاء في الوضع الصحيح، وملائمة التركيب في النظام وفقا للتوقيت والمكان المناسبين، لتتفاعل بشكل صحيح داخل النظام.(24)
الإخلال بأحد هذه الشروط يفشل الاختبار، ويؤكد عدم صلاحيته، والحقيقة التي لاحظناها بوضوح أن ميلر في اختباره لم يفعل سوى توفير جزء صغير من الشرط الأول، متمثلا في إتاحة الجزء القاعدي لإبرة الحقن والسوط، الذي لا يمثل سوى 20% من مكونات السوط، ولم يفسر وجود أجزاء المحرك الدوار الأكثر تعقيدا في السوط، أو يجد لها مثيلا بأية وظيفة ثانوية أخرى في أي نظام آخر. بالتبعية، لم يقترب قط من الشرط الثاني أو الشرط الثالث للاختبار، ليشرح كيف تزامنت أجزاء النظام أو تواجدت في نفس مكان البناء، وكيف توافقت في تنسيق بعينه لإنتاج الوظيفة المخولة، فبدلا من ذلك بادرنا باستعراضه المسرحي الهزلي، باستخدامه لأحد أجزاء مصيدة الفئران كمشبك لرابطة العنق.
ادعى ميلر زورا –حانثا باليمين الذي أقسم به قبل الإدلاء بشهادته– أن بيهي قد افترض عدم وجود وظيفة ثانوية لأجزاء النظام في تراكيب أخرى، والتي يمكنها بحسب ادعائه أن تتكيف فيما بعد لإنتاج النظام الجديد، وأطلق عليها فرضية الخيار المشترك Co–option أو التكيف المسبق Exaptation، الذي تم اصطلاحه للتعبير عن الخصائص التي تظهر في سياق وظيفي ما، قبل أن يتم استغلالها في سياق آخر، حيث يمكن لسمة معينة كانت تخدم وظيفة بعينها أن تتحول في وقت لاحق لوظيفة أخرى.
ومن الأمثلة الشهيرة للتكيف المسبق "ريش الطيور"، الذي تفترض الداروينية وجوده لتدفئة الحيوانات قديما، قبل أن يتكيف كعامل رئيس في الطيران، ومثل هذه القصص من السذاجة بمكان، حيث يمكنها أن تصلح فقط كقصص ما قبل النوم التي كانت تحكيها لنا الجدات؛ فالريش بشكله الحالي وجد خصيصا بتركيب وتوزيع ليساعد على الطيران، وكونه يحمل وظيفة أخرى تفيد في العزل والتدفئة لا يعني بالمرة أنه تكيف من هذه الوظيفة تماما، كما لا يعني وجود جيوب بمعطف التدفئة نضع بها متعلقاتنا بأن المعطف الشتوي قد صنع في البداية للاحتفاظ بالمتعلقات، ومن السذاجة بمكان ربط تطور الطيران بوجود الريش، فالخفاش يمكنه الطيران بدون ريش، كما تفعل الفراشة ويفعل اليعسوب، وإن احتاج الحيوان ليصبح طائرا فلن ينفعه زغب التدفئة في تطوير تلك الخاصية، ومثل تلك الطرق الاستدلالية تقع تحت إطار مغالطة الهجوم على رجل القش.
من ذلك المنطلق، وضع القاضي جونز في حيثيات حكمه أن بيهي كان جاهلا بآلية التكيف المسبق لتفسير تحول الوظيفة، والحقيقة أن القاضي جونز هو من يجهل تماما مثل هذه القضايا المتخصصة، فبيهي تناول هذه الحجة تفصيليا من خلال كتابه (صندوق داروين الأسود) حين تكلم عن كيفية استخدام مجموعة من القطع التي تمتلك وظائف ثانوية في نظم أخرى، كما في تطور أهداب الحركة في البكتيريا، وشرح ذلك من خلال مثاله الشهير مصيدة الفأر، حين أشار إلى فرص تكوين تلك الآلة من مهملات المرآب التي كانت تستعمل لأغراض أخرى سابقة، فقطعة خشبية يمكن استخدامها كثقالة أو لأي غرض آخر، ونابض مستخدم في ساعة قديمة مهملة، وغيرها من أجزاء تشكيل المصيدة، يقول بيهي إن الإشكالية الحقيقية تكمن في إدخال مثل هذه الأجزاء في نظام وظيفي جديد، وحاجاتها إلى سلوك مجموعة من التعديلات الملتوية بالغة التعقيد، وفيها ينعدم الدور المخول للانتخاب الطبيعي تماما، هذا هو سر فعالية الحجة، فحتى مع توافر جميع الأجزاء الضرورية المطلوبة كـ(القاعدة، النابض، عصا توقيف)، فإنه يتوجب عليها أن تتواءم مع بعضها بدقة، وإلا فالمصيدة ستكون فاشلة ولن تعمل.
أنصار التطور يدركون جيدا حجم الإشكالية، ومن ثم يجادلون في استنادهم على حجة التكيف المسبق بأن النظم الحيوية المعقدة التي تبدو غير قابلة للاختزال يتم بناؤها بطرق غير مباشرة بعملية تشبه عملية التسقيل (دعم بسقالات)، والتي يمكنها المساعدة في رفع البناء، وإيصال المكونات إلى مكانها في النظام حتى يكتمل، ومن ثم يتم إزالة هذه الأجزاء (السقالات).
الإشكالية هنا تكمن في الإجابة عن هذا التساؤل المحوري:
أي قانون طبيعي بلا هدف يمكنه أن يوجه تلك الأجزاء (بعملية التسقيل المزعومة) إلى موقعها المطلوب، في الزمن المطلوب بالتناسق المطلوب، وما هي فرص حدوث مثل هذا الحدث؟ من ناحية أخرى فالاستناد إلى مثل هذا التشبيه مرتد على صاحبه؛ فعملية البناء بالتسقيل للأبنية المعقدة بالقياس هي عملية ذكية نتاج توجيه هندسي محكم، وليست عشوائية، حيث تتطلب دقة متناهية في توجيه السقالة في مكان محدد، ونزعها في توقيت دقيق.
دعونا لا نستبق في الحكم، ونتوجه مباشرة لاختبار هذه الفرضية، نضع نموذجا افتراضيا لآلة جزيئية تتألف من مكونات البروتين (A، B، C، D)، التي تتفاعل مع بعضها لتنتج الوظيفة البيولوجية المخولة بتلك الآلة البيولوجية.
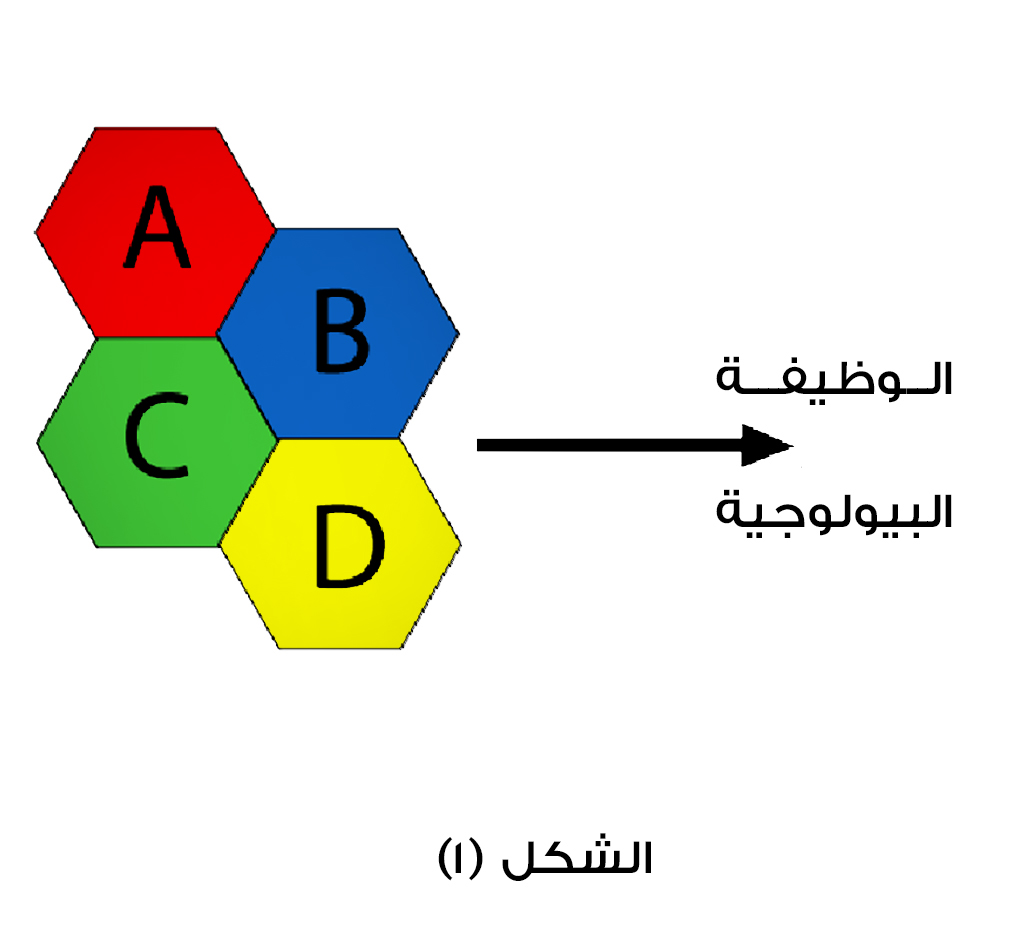
الشكل (1): مكونات (A، B، C، D)، تتفاعل لإنتاج الوظيفة البيولوجية.
إذا كان لا يمكن لهذه الوظيفة البيولوجية أن تقوم إلا بواسطة التفاعل بين المكونات السابقة، يتوجب ساعتها على أنصار التطور استدعاء فرضية التكيف المسبق لتفسير حدوثها، وفيها يفترض أن كل جزء بروتيني من الأجزاءA) ، B، C، D) كان يحمل في الأصل وظائف ثانوية سابقة.
من ثم يتفاعل مع بروتينات أخرى على مر الزمن التطوري، ويخضع النظام ككل لعمليات متكررة من التحول الوظيفي، وهذا هو جوهر التكيف المسبق، كما هو موضح في الشكل (2).
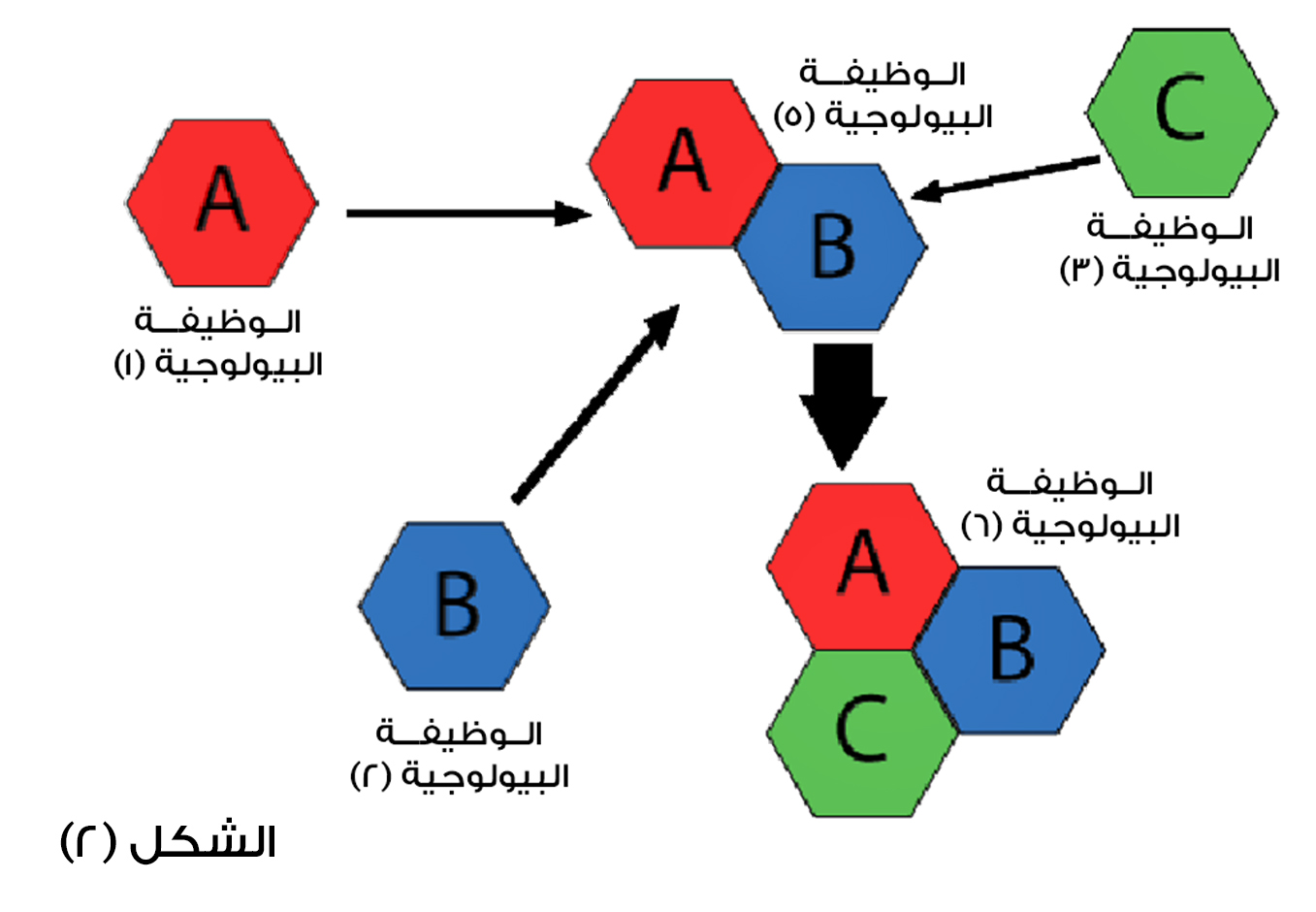
الشكل (2): المكونات تحمل وظائف ثانوية لا علاقة لها بالوظيفة الرئيسية للنظام الكلي، ويجب أن تنتج وظيفة جديدة مع كل خطوة تدريجية في سبيل تحولها إلى خطوة أخرى.
ندرك جيدا أن الآلات الجزيئية يتم بناؤها من أجزاء فردية من البروتينات، فالسوط البكتيري من السالمونيلا يتكون من 42 جزءًا من البروتين، منهاMotA وMotB (البروتينات الحركية)، FlgE (المحور).. إلخ. وتتفاعل هذه الأجزاء البروتينية مع بعضها بتكامل دقيق للغاية، يحدده أشكالها ثلاثية الأبعاد، التي يتم تعشيقها لتتآلف وتتعاون لإنتاج الوظيفة المخولة بالنظام النهائي، حيث يتوجب على بروتين يمتلك شكل به نتوءات معينة أن يقابله بروتين آخر يمتلك شقوقا متكاملة معها تماما.
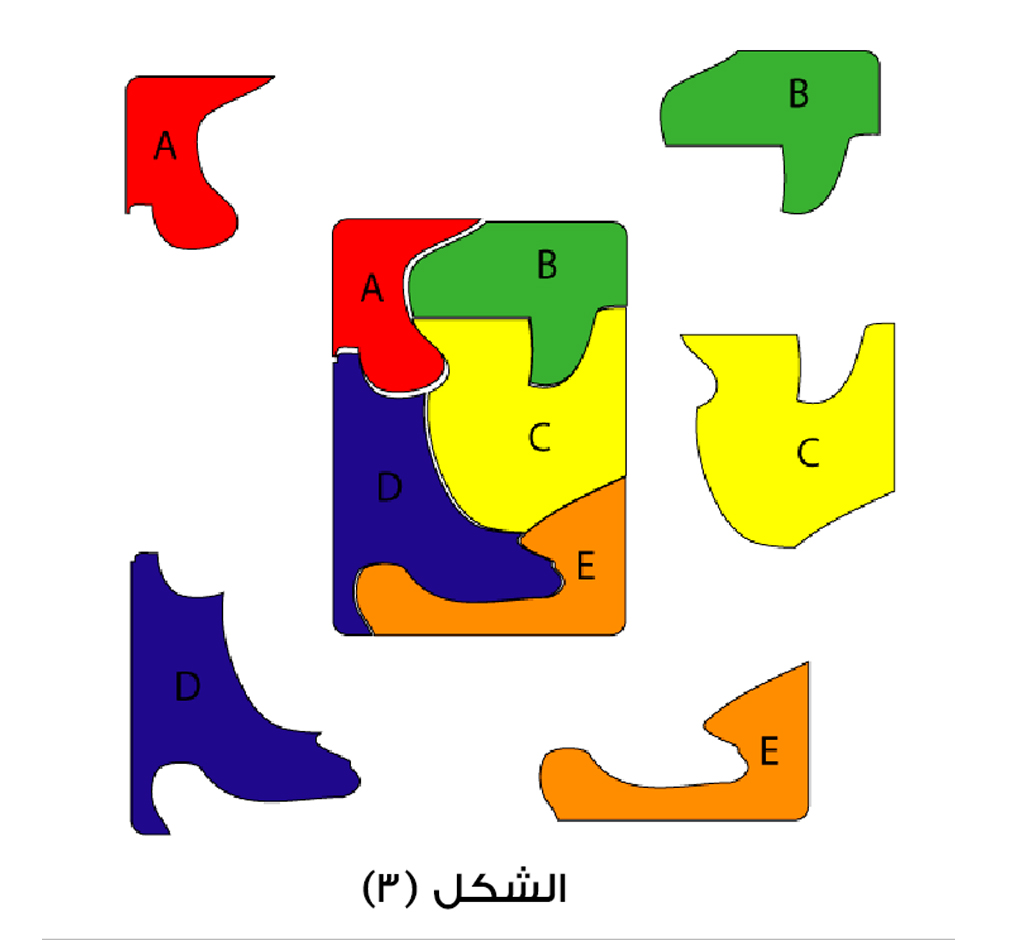
الشكل (3): نرى آلة معقدة تتكون من خمسة عناصر (بروتينات)، هي A، B، C، D، E. وهذه البروتينات هي مكملة لبعضها البعض؛ فالعنصر A هو مكمل لـ(B، C، D)، والعنصر D يعتبر مكملا لـ(A، C، E).
سيناريو التكيف المسبق يحملنا على الاعتقاد بأن أجزاء الآلة من A إلى E كانت تعمل أصلا في سياقات مختلفة، وتشكلت كل واحدة منها بصورة مستقلة عن طريق الانتخاب الطبيعي.
من ثم اجتمعت هذه الأجزاء لتتكامل في الوظيفة الجديدة، لكن في خطوة التكامل هذه ينعدم تماما دور الانتخاب الطبيعي، ويبقى العامل الوحيد المتحكم في تكامل هذه الأجزاء داخل النظام الجديد هي الصدفة وحدها، فليس هناك شيء في الانتخاب الطبيعي من شأنه الدفع نحو موائمة أشكال البروتينات لبعضها البعض، ومن المهم أن نتذكر ما شرحناه مسبقا بأن التطور غير غائي، ولا يوجد لديه بعد نظر، على عكس المهندس الذي يتحرك وفق خطة إنشائية مسبقة، فيضع السقالات في المكان الملائم ليرفع أجزاء البناء، لتتكامل في نهاية المطاف، ومن غير المعقول تماما أن نتوقع أن تلك العمليات غير الغائية يمكنها خلق هذه البروتينات فقط في الطريق الصحيح لتتوائم بدقة متناهية في الشكل والتموضع، لأداء الوظيفة الكلية للنظام.
وكما يشير كل من سكوت مينيخ Scott Minnich وستيفن ماير Stephen Meyer أنه حتى مع توافر البروتينات اللازمة لتكوين السوط البكتيري الدوار، فإنها يجب أن تترتب وفقا لتسلسل زمنى صحيح تماما كما يتم تركيب السيارة أثناء خطوات تصنيعها، ومن أجل حدوث ذلك فإنها بحاجة إلى نظام إنشائي مسبق من الشفرات والتعليمات الجينية، وأدوات التجميع والتركيب المتمثلة في آلات جزيئية من البروتينات المتخصصة لمهمة ترجمة تلك التعليمات وتنفيذها.(25)
مما سبق يتضح أن حجة أنصار التطور المركزية حول التكيف المسبق عاجزة تماما في تفسير التعقيد الحيوي ونشأته، ويتضح أيضا أن ميلر لم يقترب بأي حال من الأحوال من تفنيد التعقيد غير القابل للاختزال، بل حوّل وجهة النقد إلى مهاجمة رجل القش الذي صنعه بتحريف حجة بيهي واختزالها، من ثم وضع الاختبار في المسار الخاطئ.
والحقيقة أن فرضية الخيار المشترك بتحول الوظائف وتكيفها إلى أخرى مجرد حيلة أخرى من حيل تمرير الداروينية الشهيرة، التي أجاد ستيفن جاي جولد اختلاق الكثير منها للهروب من الإشكاليات، لكنه لم يقدم للتطور سوى نوع من تكييف المشكلة وتحويل مسارها إلى طرق جانبية ملتوية، لا تقدم أيّ حلول.
- أيهما وجد أولا؛ السوط البكتيري، أم نظام الحقن T3SS؟
كل التصورات السابقة قائمة على نقد سيناريو تطور السوط البكتيري تدريجيًّا من نظام أبسط، هو إبرة الحقن T3SS كما تنبأت الداروينية. فماذا لو علمنا أن وجهات الفحص تصب في صالح تصور معاكس تماما؛ حيث تفترض الدراسات أن السوط البكتيري كان سابقا لظهور إبر الحقن، وأن البكتيريا لابد أنها احتاجت للدفع والحركة قبل أن تحتاج أدوات الافتراس التي تستخدم لمهاجمة خلايا حقيقات النواة Eukaryotes، والتي تطورت في وقت لاحق. ومن الأسباب الداعمة أيضا لتلك الفكرة هي أنه قد تم العثور على T3SS في نطاق ضيق من البكتيريا، في حين وجدت الأسواط البكتيرية على نطاق واسع في المجموعات البكتيرية المختلفة، وهو ما يعني أنها قد نشأت في وقت مبكر عن سابقتها.
هاورد أوكمانHoward Ochmanالمتخصص في الكيمياء الحيوية في جامعة أريزونا، يشير إلى تطور T3SSمن السوط وليس العكس، فكيف يمكن للداروينية تخطي ذلك الأمر؟(26)
- فجوة معرفية، أم دليلي إيجابي؟
مما لا شك فيه أن فكرة عداء العلم للدين في الغرب تعود بقسم كبير منها إلى ذلك الإرث الكنسي اللاهوتي القديم المغرق في السطحية، الذي وضع برهانا خاطئا على وجود الله ينحصر في نوع من المعجزات، مرتبط بما لا نستطيع تفسيره، وكان تبني تلك الفكرة المهينة بأن الله لا يتواجد إلا في الجزء غير المفسر من العالم بمثابة الحبل الذي لفه اللاهوتيون حول أعناقهم، ومن ثم كانت الفرصة الذهبية لتنهي العلمانية المهمة بسهولة، وتركل الكرسي من تحت أقدام اللاهوتيين.
وعليه، لا يمكننا تشديد اللوم على المادية في اصطلاحها المشهور (إله الفجوات المعرفية)، لكن في المقابل نجد أنه من الأهمية تصحيح المفاهيم، وإعادة توجيه التوصيف إلى مساره، فتغيير الفكرة السائدة حول مرض الصرع ومعرفة أسبابه، وتصحيح الفكرة التي سادت حوله باعتباره تلبس بروح شريرة، وأن علاجه يكمن في استجلاب معجزات إلهية بالصلوات، في الحقيقة لا يعتبر هدما للإله وانزواء لدوره في مقابل الحل المادي، بقدر ما يمكن اعتباره هدما للفكرة الخاطئة التي تبناها اللاهوتيون حول دور الله وتوظيفه بالخطأ، والذي اعتمد على الالتجاء إلى نوع من التواكل وعدم السعي لمعرفة الأسباب وتتبعها، لذا ظلت العلمانية تكسب جولات زائفة بتراجع دور الإله بذلك المفهوم، لكنها في الحقيقة لم تجابه إلا (رجل القش) الذي ساهم اللاهوتيون أنفسهم في خلقه هذه المرة. فنجاح المادية في تفسير كيفية عمل الآلة المعقدة التي كانت تبدو من قبل ذلك للبعض كالسحر والمعجزة، لا يمكنه أن ينفي وجود صانع لتلك الآلة ويرجع الأمر برمته إلى القوانين الطبيعية، مدعيا قدرة تلك القوانين على صنعها، بل على النقيض من ذلك يجب أن يزيد يقيننا في قدرة الصانع وبراعته، ويؤكد قناعتنا بحتمية التصميم الحكيم.
من جهة أخرى، فإن الفجوات المعرفية المحيطة بقضية ما –على فرض صحتها– يتوجب عليها أن تعطي لدى الباحث المحقق انطباعا من الحيادية أو اللاأدرية بخصوص تلك القضية، وعدم استباق الأحكام حولها، لكن في الوقت الذي يدعي فيه أنصار المادية استعانة المعارضين بإله الفجوات لسد تلك الثغرات المعرفية، فإنهم في المقابل يسدونها بإله آخر؛ هو (الصدفة العشوائية)، وينصبون التفسير المادي الطبيعي ليقعوا تحت نفس إطار الاتهام سد الثغرات بإله الفجوات.
حتى لا نتشعب كثيرا في تفاصيل فرعية، يمكن للإشارة السابقة أن تكون كافية لإلقاء الضوء على ذلك الادعاء الشائع، الذي يروج له أنصار المادية بأن التعقيد الحيوي غير الاختزالي لا يمكن اعتباره برهانا إيجابيا داعما للتصميم، بقدر ما هو برهان سلبي مبني على استغلال فجوات معرفية متعلقة بجهل آليات نشوء هذه الأعضاء الحيوية، التي يمكن سدها في المستقبل.
لكن الحقيقة غير ذلك، فهذه النظم تبدي علامات التصميم الحكيم التي يستحيل تفسيرها من خلال عمليات طبيعية عشوائية، ومن خلال قياس بسيط يمكننا إدراك تلك الحقيقة، فلو وجد شخص ما كهفًا حجريًّا مصقولا وسط الجبال، سيستنتج أنه تم صنعه بفعل مصمم، لكنه أيضا لن يبرر بنفس الادعاء إذا وجد قطعا صخرية عشوائية الشكل ومن نفس الحجم.
- التعقيد المتخصص
تتميز الحياة بتعقيد تفشل في مضاهاته أيّة معقدات غير حية، ذلك أنها لا تتعدى كونها معقدات عشوائية ترتبط خلالها الوحدات الصغيرة من خلال روابط كيميائية كأحجار الكريستال.
نرى في نماذج التتابع الجزيئي في الجزيئات البيولوجية الوظيفية –مثل النظام المعلوماتي للحياة المعروف بالحمض النووي DNA– والتي أسماها ويليام ديمبسكي التعقيد المتخصص Specified Complexity، ويعرفه كعلامة واضحة من علامات التصميم الذكي بقوله:
"عندما يبدي شيء ما تعقيدًا متخصصًا، أي عندما يكون معقدًا ومتخصصًا بنفس الوقت، فإننا نستطيع أن نقول أنه قد أُنتج من قبل مسبب ذكي، عوضًا عن القول بأنه كان نتيجة للعمليات الطبيعية".(27)
فالتعقيد المتخصص نظام لا يكتفي بالتعقيد العشوائي، بل بتخصص التعقيد لأداء أدوار ومهام محددة، ويستخدم ديمبسكي المثال التالي:
• الحرف الأبجدي هو متخصص دون كونه تعقيدا.
• جملة طويلة من الأحرف العشوائية هي تعقيد دون كونه متخصصا.
• قصيدة لشكسبير هي تعقيد متخصص.
ويطرح ديمبسكي نموذجا رياضيًّا لتقنين التعقيد المتخصص، في إطار ما قام بتعريفه بـ(حد الاحتمال الكوني)؛ فلو كان عدد الجسيمات الأولية داخل الكون هو 8010، وعدد العمليات الفيزيائية التي يمكن حدوثها في الثانية الواحدة هي 4510، وعمر الكون منذ الانفجار الكبير بالثواني هو 2510، فحاصل ضرب العوامل السابقة يمكن أن يعطينا الحد الأقصى للأحداث التي يمكن أن تكون قد حدثت للجسيمات الأولية، منذ الانفجار الكبير حتي وقتنا هذا، لذلك فإن أقل احتمالية حدوث حدث عشوائي خلال تاريخ الكون هو واحد من (15010). أما الأحداث التي تكون قيمة احتمالها أقل من هذه القيمة فمن غير الممكن حدوثها بكوننا بشكل عشوائي، ومنه يعرف ديمبسكي المعلومات المعقدة المتخصصة بأنها أي شيء احتمال حدوثه في الطبيعة أقل من 1 من 15010.(28)
بالعودة إلى السؤال الذي طرحناه في التمهيد حول سر وجود النظام الحيوي، يمكننا الآن أن نكوِّنَ تصورًا للإجابة عنه، في ظل التقدم العلمي في مجال (البيولوجيا الجزيئية)، الذي منحنا نظرة أكثر عمقا عن سيرورته، وكشف لنا –مع ذلك التقدم– أن الكثير من التفاعلات المهمة بين مكونات أي كائن متعض لا تتم على المستوى الفيزيوكيميائي، بل على مستوى تكاملي أرقى منه ومتسيد عليه، هو البرنامج المعلوماتى المتمثل في الحمض النووى DNA، الذي يتواجد داخل نواة كل خلية حية، وهو بمثابة أبجدية مكونة من أربعة أحرف، تحمل المعلومات ذات التعقيد المتخصص، تماما مثل الجمل العربية أو برامج الكمبيوتر، والتي لا يمكن تفسيرها بكيمياء الحبر أو فيزياء المغناطيسية، لكنها ترجع بالضرورة إلى التصميم الحكيم. هذا البرنامج يمكن اعتباره القاسم المشترك بين جميع الكائنات الحية من البكتيريا إلى الإنسان، وهو ما يقوم بوظيفة توجيه الأجزاء إلى الترابط في تلك النظم الحيوية على نحو دقيق وقيادتها، ويحمل مخططات بناء الكائن الحي ووظائفه بكل تفاصيله المدهشة.
يشير (پول ديڤيز Paul Davies) عالم الفيزياء النظرية والبيولوجيا الفلكية إلى تلك الحقيقة بقوله: "بإرجاء الحياة إلى قوانين الفيزياء أو الكيمياء نراها تبدو مثل السحر، إنها تتصرف بطرق غير عادية لا مثيل لها في أي نظام فيزيائي أو كيميائي آخر؛ فهي تحمل خصائص نابضة بالحياة تتميز بالاستقلالية والقدرة على التكيف، والسلوك الموجه نحو الأهداف، وتسخير التفاعلات الكيميائية لتمرير أجندة مبرمجة مسبقا، بدلا من أن تكون عبدا لتلك التفاعلات".(29)
في نفس السياق يقول: "نعرف الآن أن سر الحياة لا يكمن في المكونات الكيميائية على هذا النحو، بل في البنية المنطقية والترتيب التنظيمي للجزيئات، فالحياة هي نظام معالجة المعلومات، وبرنامج الخلية الحية هو السر الحقيقي، وليست الأجهزة، لكن من أين أتى البرنامج؟ كيف لذرات غبية بشكل عفوي كتابة البرامج الخاصة بها؟ لا أحد يعرف".(30)
هذه الفجوة المعرفية التي يدعيها أنصار المنهج المادي حول مصدر البرامج والمعلومات الحيوية هي في حقيقتها ليست إلا فجوة مصطنعة يتم الترويج لها، بغرض الهروب مما تؤول إليه التحليلات المنهجية من نتائج في هذا الصدد.
على النقيض، نجد أن الكثير من الأبحاث المعنية تؤكد حتمية تصدر التصميم لخلق مثل هذه البرامج المعلوماتية الحيوية، ففي بحث بعنوان (في البدء كانت المعلومات In the Beginning Was Information) لجيت فيرنر GittWerner ، أستاذ الفيزياء ومدير قسم معالجة المعلومات في معهد الفيزياء والتكنولوجيا براونشفايج، يخلص إلى هذه النتيجة بقوله: "نظام الترميز يستلزم دائما عملية عقلية. النهج الفيزيائي لا يمكنه أن ينتج رموز المعلومات. تُظهر جميع التجارب أن كل قطعة من المعلومات الإبداعية تمثل بعض الجهد العقلي".(31)
في نفس السياق يعترف كل من ليستر L. Lester وبولين R. Bohlin بتلك الحقيقة: "الحمض النووي هو رمز المعلومات... الاستنتاج القطعي هو أن هذه المعلومات لا يمكن أن تنشأ تلقائيًّا من خلال عمليات آلية. الذكاء ضرورة في الأصل لأى رمز معلوماتي، بما في ذلك الشفرة الوراثية".(32)
- خاتمة
من خلال التحليل التتابعي السابق، نخلص إلى النقاط الآتية:
• الكائن الحي ليس تراكمات مجردة لمجموعة من المفردات، بل نظام دقيق متكامل لا يقبل الاختزال أو التطور التدريجي.
• يتحكم في سيرورة هذا النظام (برنامج معلوماتي) إنشائي وتشغيلي مسبق، يمثل نوع من التعقيد المتخصص.
• البرنامج المعلوماتي يستحيل تفسير وجوده بالنهج الفيزيوكيميائي والقوانين الطبيعية، فهو نتاج حتمي للتصميم، كما أثبتت التجارب والدراسات في هذا الشأن.
دمتم بود، وإلى لقاء آخر بإذن الله مع فصل جديد من فصول (إعادة المحاكمة).
المراجع:
(1) "If it could be demonstrated that any complex organ existed, which could not possibly have been formed by numerous, successive, slight modifications, my theory would absolutely break down."
Charles Darwin, "The Origin of Species", Harvard University Press, 1964, p. 189.
(2) Ernst Mayr, "This Is Biology: The Science of the Living World", Harvard University Press, 1998.
(3) David Quammen, "The Reluctant Mr. Darwin: An Intimate Portrait of Charles Darwin and the Making of His Theory of Evolution (Great Discoveries)", W. W. Norton 2006.
(4) Charles Darwin, "The Origin of Species", Harvard University Press, 1964, p. 190.
(5) "the crucial importance of this requirement to the theory of evolution was fully understood by Darwin, who stated that, in searching for the gradations through which an organ in any species has been perfected, we ought to look at its lineal progenitors. Indeed we ought; though he himself could not do so. It is deceptive to the reader to create a seriation beginning with eye spots as seen in unicellular organisms and call them, as does Duke–Elder (1958), the earliest stage of evolution."
Cousins, F.W., "The Anatomy of Evolution, Duffett Publications", London, p. 125, 2003.
(6) Atsushi Ogura. et al, "Comparative Analysis of Gene Expression for Convergent Evolution of Camera Eye Between Octopus and Human", Genome Res. 2004.
genome.cshlp.org/content/14/8/1555.short
(7) Fernald, R.D., "The evolution of eyes", Brain, Behavior and Evolution 50 (4):253, 1997.
(8) Frank Salisbury, "Doubts About the Modern Synthetic Theory of Evolution", American Biology Teacher, September 1971, p. 338
(9) Fernald, R.D., "Eyes: variety, development and evolution", Brain, Behavior and Evolution 64(3):141–147, 2004; p. 1917.
(10) Croft, L.R., "The Last Dinosaurs", Elmwood Books, Chorley, Lancashire, p. 57, 1982.
(11) Land, M.F. and Nilsson, D.–E., "Animal Eyes", Oxford University Press, New York, p. 1, 2005.
(12) Duke–Elder, S.S., "System of Ophthalmology. Volume 1: The Eye in Evolution", The C.V. Mosby Company, St. Louis, p. 237–238
(13) Breidach, O. and Kutsch, W., "The Nervous Systems of Invertebrates: An Evolutionary and Comparative Approach." With a coda written by T.H. Bullock, 1995.
(14) Fernald, R.D., "Casting a genetic light on the evolution of eyes", Science 313:1914–1918, 2006; p. 1914.
(15) Turner, J.S., "The Tinker's Accomplice: How Design Emerges from Life Itself", Harvard University Press, Cambridge, MA, p. 161, 2007.
(16) "Have you ever seen a mutation simultaneously affecting two separate components of the body and producing structures that fit one another precisely? ... have you ever beheld three, four or five simultaneous mutations with matching structures producing coordinating effects? ... These are vital questions that demand an answer. There is no way of getting around them, or evading the issue. Every biologist who wants to know the truth must answer them, or be considered a sectarian and not a scientist. In science there is no "cause" to be defended, only truth to be discovered. How many chance occurrences would it take to build this extraordinary creature [Myrmelion formicarius]'?"
Grassé, P.P., "Evolution of Living Organisms", Academic Press, New York, NY, p. 163, 1977.
(17) Joseph A. Kuhn, "Dissecting Darwinism", Proc (Bayl Univ Med Cent). Jan 2012; 25(1): 41–47.
(18) Michael Behe, "Darwin's Black Box: The Biochemical Challengue to Evolution", 10th ed. (2006) Free Press, New York.
(19) KITZMILLER, et al. v. DOVER AREA SCHOOL DISTRICT, et al. – Case No. 04cv2688 – Middle District of Pennsylvania Court.
(20) عادل مصطفى، المغالطات المنطقية، المجلس الأعلى للثقافة 2007، ص163.
(21) Dr. Kenneth Miller Testimony, Day 1, PM Session, page 16.
(22) Casey Luskin, "Do Car Engines Run on Lugnuts? A Response to Ken Miller & Judge Jones's Straw Tests of Irreducible Complexity for the Bacterial Flagellum.", CSC – Discovery Institute, April 19, 2006. discovery.org/a/3718
(23) Dembski, Rebuttal to Reports by Opposing Expert Witnesses , p 52.
www.designinference.com/documents/2005.09.Expert_Rebuttal_Dembski.pdf
(24) Angus Menuge, "Agents Under Fire: Materialism and the Rationality of Science", p 104–105, Rowman & Littlefield, 2004.
(25) Scott A. Minnich y Stephen C. Meyer, "Genetic Analysis of coordinate flagellar and type III regulatory circuits in pathogenic bacteria", Discovery Institute p 8.
www.discovery.org/scripts/viewDB/filesDB–download.php?id=389
(26) Dan Jones, "Uncovering the evolution of the bacterial flagellum," New Scientist (Feb 16, 2008).
(27) Dembski. "Intelligent Design", p. 47 www.designinference.com/documents/2003.08.Encyc_of_Relig.htm
(28) Dembski, (ed.) "Mere Creation: Science, Faith & Intelligent Design.", Downers Grove, IL: InterVarsity Press, 1998, 209–213.
(29) "To a physicist or chemist life seems like 'magic matter,'" Davies explained. "It behaves in extraordinary ways that are unmatched in any other complex physical or chemical system. Such lifelike properties include autonomy, adaptability and goal–oriented behavior –– the ability to harness chemical reactions to enact a pre–programmed agenda, rather than being a slave to those reactions."
Skip Derra, "ASU researchers propose new way to look at the dawn of life", asu news Posted: December 12, 2012.
www.asunews.asu.edu/20121212_dawnoflife
(30) (a) Paul Davies, "The Origin of Life: Fifth Miracle", Penguin UK.
(b) New way to look at dawn of life, www.sciencedaily.com/releases/2012/12/121212205918.htm
(c) Origin of Life Needs a Rethink, Scientists Argue, www.livescience.com/25453–life–origin–reframed.html
(31) "A coding system always entails a nonmaterial intellectual process. A physical matter cannot produce an information code. All experiences show that every piece of creative information represents some mental effort and can be traced to a personal idea–giver who exercised his own free–will, and who is endowed with an intelligent mind."
Werner Gitt, "In the Beginning Was Information", CLV, Bielefeld, Germany, pp. 107, 141.
(32) "DNA is an information code. . . . The overwhelming conclusion is that information does not and cannot arise spontaneously by mechanistic processes. Intelligence is a necessity in the origin of any informational code, including the genetic code, no matter how much time is given."
L. Lester and R. Bohlin, "The Natural Limits to Biological Change", (Dallas, TX: Probe Books, 1989), p. 157